|
Current location: Home > Physical Processes Model > Basic Concepts |
|
Basic Concepts Related to Primary and Secondary Drivers Jump down this page to: This section provides general background information to the reader on each of the primary and secondary drivers included in the Physical and Chemical Processes Model. This includes information on basic physical and chemical principles and a discussion of interactions among the primary and secondary drivers. Information provided in the model itself is specific to each of the river reaches. The primary drivers affecting DO concentrations are described below. These drivers are the same in each of the three reaches in the study area. Each of the primary drivers is first defined, and where available, sources of the driver are described, as well as how each driver is measured. In addition, links are provided to the discussions of how the primary drivers affect each other and to descriptions of the secondary drivers that affect the primary drivers. Definition Imported DO is DO that is contributed to each reach of the San Joaquin River in water from upstream, tributaries, return flows, and other flow sources. Imported DO contributes to the initial DO concentration of water flowing into each reach. Definition Reaeration, a natural physical process, is the net rate of transfer of oxygen from the atmosphere to a body of water with a free surface (U.S. Environmental Protection Agency 1999a). This transfer occurs at the air/water interface. The transfer of DO from the atmosphere to the surface of a waterbody is controlled by the transfer rate and the DO deficit. Reaeration is at its minimum, zero, when the DO concentration is at saturation and is at its maximum when the DO concentration is zero. Reaeration increases with surface area. Reaeration is therefore greater in the DWSC, where the surface width is 600 feet, than in the rest of the San Joaquin River. The transfer rate depends on the turbulence of water near the water surface, which is generally estimated from the water velocity and depth (or other hydraulic parameters), as well as the wind speed. Water depth may influence turbulence at the surface and is included in most regression equations used for estimating the transfer velocity. The rate of change in DO always decreases as the mixed depth increases (Jones & Stokes 2003). Stratification limits the mixed depth that is exposed to surface reaeration. When a water body is stratified, with a warmer layer isolated near the surface layer, reaeration is also limited to the surface. The surface mixed depth increases when wind speeds are higher and at night from cooling. This greater mixed depth allows a larger fraction of the water volume to be exposed to reaeration, so a greater mass of oxygen is transferred to the water body during periods of higher wind speed (Jones & Stokes 2003). Measuring Reaeration Rates Reaeration is calculated as: reaeration [g/m2/day] = transfer rate [m/day]• DO deficit [mg/L]. The general mass balance equation for oxygen gas transfer from the atmosphere to a mixed volume of water exposed to the atmosphere is: V • dC/dt = KL • A • (Cs– C)
This relationship is often written as: dC/dt = K2 • (Cs– C)
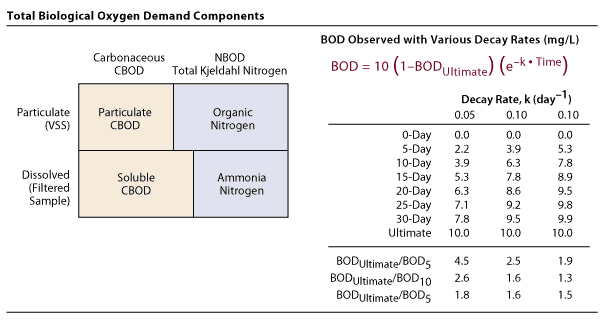
Interactions with Other Primary Drivers Reaeration interacts with the following primary drivers: residence time, photosynthesis, and imported DO. Secondary Drivers That Affect Reaeration The following secondary drivers affect reaeration: water temperature, flow velocity, and channel geometry. Definition Aquatic vascular plants, attached algae (periphytes), and free-floating algae (phytoplankton) produce oxygen through photosynthesis, which contributes to DO concentrations in the water. The simplified chemical reaction that produces oxygen through photosynthesis (Tchobanoglous and Schroeder 1985) is: CO2 + H2O + Nutrients (in the presence of light) —> CH2O [new algal cells] + O2 This equation suggests that for each gram of algal biomass produced, about 2.67 grams of O2 will be produced. However, algal biomass is about 40% carbon, so 1 gram of biomass will be associated with about 1 gram of oxygen. Therefore, for each 1 mg/L of DO produced during the day in the San Joaquin River at Mossdale or near the surface of the DWSC, an equivalent algal biomass must have been produced during photosynthesis. Interactions with Other Primary Drivers Photosynthesis interacts with the following primary drivers: reaeration and residence time. Secondary Drivers That Affect Photosynthesis The following secondary drivers affect photosynthesis: algal biomass, turbidity, water temperature, and nutrients. The amount of sunlight available for photosynthesis is determined by diurnal and seasonal variations in light intensity and by weather-related factors, such as the presence and density of cloud cover. In some cases, vegetative cover from river banks also affects the amount of sunlight reaching the water surface. The amount of sunlight available for photosynthesis is governed by factors that are not controllable (except for shading from riparian trees) and, therefore, this secondary driver is not discussed in the reach-specific sections. Maximum rates of algal photosynthesis occur at 2545% of the light intensity available at the DWSC water surface (Lehman et al. 2001). Definition BOD is the consumption (or decrease) of DO, caused either by microorganisms as they decompose organic material or by chemical oxidation. BOD measures the potential of certain materials to decrease DO concentrations as these materials are converted to other substances. The two types of BOD are described below. Episodes of low DO concentrations in the DWSC are generally caused by high concentrations of oxygen-demanding substances that decay in the DWSC during periods of relatively long residence time. Carbonaceous Biochemical Oxygen Demand BOD related to microorganism is called CBOD. The source material for CBOD is organic matter. CBOD results when oxygen is consumed by microorganisms in converting organic material into CO2, H20, nutrients, energy, and new cells (Lee and Jones-Lee 2003). Algal cells contain organic chemicals that consume oxygen during decomposition. The following chemical equation provides estimates of the quantity of CBOD that is produced during the decomposition of an algal cell (Litton 2003). C106H263O110N16P (algae) + 138 O2 —> Nitrogenous Biochemical Oxygen Demand BOD related to chemical oxidation is called NBOD. The source materials for NBOD include organic matter that decays to ammonia, and ammonia entering the San Joaquin River through wastewater and stormwater systems. Nitrification, the process of oxidizing ammonia to nitrates by microorganisms, requires almost 5 mg/L of DO (NBOD) for every mg/L of ammonia that is oxidized. Sources of BOD in the San Joaquin River Known and potential sources of BOD in the study area include algae, aquatic plants, dead plant and animal matter, and other soluble and insoluble organic materials. It is difficult to determine the contribution of specific organic materials to BOD, but it is known that each compound is subject to a specific rate of biochemical reaction that consumes DO (Lee and Jones-Lee 2003). The current body of information indicates that algae and algal detritus are the primary sources of BOD in the DWSC (Lee and Jones-Lee 2003). Each 1 mg/L of algae yields a theoretical total oxygen demand of 1.2 mg/L, which represents the sum of CBOD and NBOD. The CBOD from the decomposition of 1 mg/L of algal biomass is 0.9 mg/L, or 75% of the total oxygen demand, while the NBOD is 0.3 mg/L, or 25% of the total oxygen demand (Litton 2003). The algae pigments chlorophyll a and pheophytin are used to represent the concentrations of living and dead algae, respectively, in a water column. Therefore, the pheophytin concentrations indicate the quantity of dead algae that might be a source of CBOD. However, the chlorophyll a and pheophytin content of algal biomass is somewhat variable. There are two sources of NBOD in the study area. The first source is organic nitrogen (the nitrogen contained in organic material) that is converted by microorganisms to ammonia (NH3). This process is called ammonification. In the presence of oxygen, other microorganisms convert the ammonia into nitrites (NO2-) and nitrates (NO3-), consuming DO in the process. This process is called nitrification. The second source of NBOD is direct inputs of ammonia from wastewater treatment plants and other sources. Ammonia and organic nitrogen compounds vary in their NBOD exertion rates, which are also dependent on the number and species of microorganisms acting on the material (Lee and Jones-Lee 2003). Measuring BOD Organic materials create both CBOD and NBOD through different biochemical pathways, making it difficult to separate out the potential oxygen demand of each pathway. Measurement of BOD is usually accomplished through laboratory experiments with water samples from the target water body. BOD and CBOD are directly measured, and NBOD is inferred by subtracting the CBOD from the BOD. However, it should be noted that the nitrification-inhibitor method that has been used to measure CBOD may yield unreliable results because the inhibitor also inhibits the growth of some bacteria that contribute to CBOD (Lee and Jones-Lee 2003).
Interactions with Other Primary Drivers BOD interacts with the following primary drivers: residence time and SOD. Secondary Drivers That Affect Biochemical Oxygen Demand The following secondary drivers affect BOD: imported BOD concentrations, CBOD concentrations, NBOD concentrations, and water temperature. Definition SOD is the overall demand for DO from the water column that is exerted by the combination of biological, biochemical, and chemical processes at the sediment-water interface. The primary sources of SOD are anaerobic (low-oxygen) chemical compounds in the riverbed sediments and particulate BOD (including algae and other sources of organic matter) that settle out of the water column. SOD is generally composed of biological respiration from benthic organisms and the biochemical (i.e., bacterial) decay processes in the top layer of deposited sediments, together with the release of oxygen-demanding (i.e., reduced) anaerobic chemicals, such as iron, manganese, sulfide, and ammonia. These soluble chemicals are released into the water and exert a relatively rapid (i.e., it occurs on a timescale of hours) oxygen demand as the reduced chemicals are oxidized. Some oxidation processes, such as nitrification of ammonia to nitrate, require bacteria and may be slower (i.e., days). Sources of SOD in the San Joaquin River SOD is caused by the consumption of oxygen by bacteria as they respire and decompose algae and other organic materials that have settled to the San Joaquin River channel bottom (Lee and Jones-Lee 2000). Nitrification of ammonia present in the water column near the sediments can be an additional source of SOD (Lee and Jones-Lee 2003). Another potential source of SOD are ferric iron and reduced sulfur compounds (under anaerobic conditions) in the bed sediments (Lee and Jones-Lee 2000). If these reduced forms of iron and sulfur (i.e., ferrous iron and sulfides) are exposed to DO, they rapidly undergo oxidation reactions and, in the process, exert a high oxygen demand for a short time (Lee and Jones-Lee 2000). Only sediments on the channel bottom exert an SOD (Chen and Tsai 2002). Particulate forms of BOD may contribute to the SOD through settling. Algae and suspended particles entering the DWSC from the San Joaquin River can settle out of the water column at an average rate of approximately 1 m (3.28 feet) per hour, creating a source of SOD (Litton 2003). Resuspension and mixing may reduce the net settling rate. The potential sources of organic particles entering the study area include agricultural return flows, domestic and industrial wastewater sources, and stormwater runoff (Lee and Jones-Lee 2000). Upstream of the DWSC, where the channel is relatively narrow and shallow compared to the DWSC, the San Joaquin River is turbulent and has sufficient flow velocities to suspend algae and other organic particulates in the water. However, as the river flows into the DWSC, the flow velocities dramatically decline because of the greater width and depth of the DWSC. This velocity reduction causes some of the suspended load to settle out and contribute to SOD (Chen and Tsai 2002). Particulate matter generally decreases with distance downstream in the DWSC, and there is a corresponding decrease in the quantity of SOD (Litton 2003). Measuring SOD SOD produces an oxygen depletion rate in the water that is dependent on the:
The DO decline is calculated as: DO decline [mg/L/day] = (SOD rate [g/m2/day])/(depth [m]) • (residence time [days]) If the SOD is uniform along a river reach or in the DWSC, the residence time can be used to estimate the DO decline as the water flows across the sediments during each day of travel. For river segments with a depth of about 1 m (3.3 feet), a SOD of 1 g/m2/day reduces the DO concentration by 1 mg/L each day. Often, however, the rate of reaeration (g/m2/day) across the nearly equal area of water surface of a river is greater than the SOD rate, so there is no measured decline in DO along the river reach. In a river with a depth of 1 m, the DO decline is equal to the SOD rate times the residence time (which may be relatively short). However, in the DWSC, where the depth is about 10 m, the DO decline is only about 10% of the SOD rate times the residence time. Consequently, substantial DO decline may occur only when the residence time in the DWSC is long (i.e., during low-flow events). Practically speaking, there is no direct way to distinguish between DO declines caused by BOD (i.e., algal respiration, bacterial decay, or nitrification) or SOD. The conceptual model includes both sources of oxygen demands, although the relative magnitude of the sediment and suspended sources is uncertain. In addition, the bottom materials that produce the measured SOD are organic particulates that have settled from the water column. Interactions with Other Primary Drivers SOD interacts with the following primary drivers: residence time and BOD. Secondary Drivers That Affect Sediment Oxygen Demand The following secondary drivers affect SOD: particulate SOD concentration, iron and sulfate compound concentrations, and water temperature. Definition Residence time is the amount of time that water remains in a water body. Generally, increasing flow volume decreases residence time by increasing flow velocity in the channel. Channel geometry (i.e., width, depth) determines residence time by affecting the velocity with which a given volume of water passes through a channel segment. In the DWSC, residence time is controlled by its geometry (depth and width), and inflows from the San Joaquin River. Tidal flows increase the movement of water in the DWSC but do not change the average residence time. Particulate materials that settle to the channel bottom have a longer residence time than those in the water (i.e., dissolved substances). Measuring Residence Time Residence time can be calculated from the flow and volume: time (days) = 0.5 • volume (af) / river flow (cfs) Interactions with Other Primary Drivers Residence time interacts with the following primary drivers: BOD, SOD, reaeration, and photosynthesis. Secondary Drivers That Affect Residence Time The following secondary drivers affect BOD: flow and channel geometry. Relationships between Primary Drivers Although the graphic at the top of this page displays each of the primary drivers as acting independently on DO concentrations, in reality the primary drivers interact and affect each other as well. These relationships between the primary drivers are described below. Relationship between Residence Time and BOD Residence time affects BOD concentrations (and DO concentrations) by controlling the period over which BOD substances can exert an oxygen demand in a river reach. The longer the residence time, the more BOD is created by decay and the more DO concentration declines. Residence time also affects BOD concentrations because particulate forms of BOD, such as dead algae, settle slowly from the water column, reducing the BOD concentration. A longer residence time allows more of the particulate BOD to settle out and contribute to SOD. Relationship between Reaeration and BOD and SOD BOD and SOD concentrations can affect the rate of reaeration by reducing the concentration of DO in the water column and increasing the DO deficit. Relationship between Residence Time and SOD Residence time affects SOD concentrations because particulate BOD, such as caused by dead algae, settles from the water column and contributes to SOD. As residence time increases, the amount of particulate BOD that settles out also increases, which in turn contributes to an increase in SOD. Relationship between Residence Time and Reaeration Residence time affects reaeration by controlling the amount of time that reaeration is allowed to act on the water moving through the DWSC. The greater the residence time, the greater amount of reaeration that occurs, until saturation is reached. Relationship between Residence Time and Photosynthesis Residence time affects the amount of DO produced through photosynthesis by affecting the growth of algal biomass. The longer the residence time, the more time that algae have to grow and produce oxygen. Relationship between Photosynthesis and Reaeration Photosynthesis by aquatic plants contributes to DO concentrations and, as DO concentrations provided by photosynthesis increase toward saturation levels, the rate of reaeration decreases (Jones & Stokes 2003). The maximum long-term DO concentration is the saturated DO concentration in the San Joaquin River or the DWSC. Relationship between Imported DO Concentration and Reaeration A higher imported DO concentration also reduces the DO deficit and results in decreased reaeration (see Reaeration). Secondary Drivers Affecting DO Water temperature affects the following primary drivers: reaeration, photosynthesis, BOD, and SOD. Water Temperature Effects on Reaeration DO concentrations are controlled by Henrys law, which indicates that a small fraction of a gas will dissolve in a liquid. The maximum concentration of DO in freshwater is calculated from Henrys constant, which is a function of temperature. Increasing temperature tends to reduce DO concentrations by reducing oxygens solubility in water. The table below shows the saturation DO concentrations in freshwater for a range of water temperatures (American Public Health Association 1989). During the summer, with a water temperature of 24°C (75°F), the DO saturation concentration is about 8.5 mg/L. During the winter, with a water temperature of 10°C (50°F), the DO saturation concentration is about 11.3 mg/L. This physical chemistry principle explains why DO concentrations are generally higher in the winter and lower in the summer in the San Joaquin River and the DWSC.
Stratification (i.e., surface heating) can produce a shallow surface layer and limit reaeration of water below the surface layer. Stratification is more likely to occur during the afternoon of warm days (i.e., high solar radiation) in the DWSC. Water Temperature Effects on Photosynthesis and Biochemical Oxygen Demand Water temperature affects photosynthesis (via algal biomass) and BOD concentrations in several ways. Water temperature affects the rate that oxygen is generated through photosynthesis by affecting the growth rate of algae and other aquatic plants. As water temperatures in the San Joaquin River rise, algal growth rates increase, as do the rates of DO-depleting reactions (i.e., decay and respiration) (Lee and Jones-Lee 2003). The assumed effect of temperature on algal growth and decay is: growth/decay rate = (growth/decay rate at 20°C) • 1.05T-20
The seasonal effect of temperature on the rates is strong, but the magnitude of the effect is uncertain because no direct measurements have been made of these process rates at varying temperatures. One author has found that first-order decay constants (measured at 20 ºC) for BOD at Mossdale in the San Joaquin River and in the DWSC are approximately 0.11 per day (Litton 2001a). Some of the year-to-year variations in DO depletion in the DWSC may be related to temperature differences, which influence algal growth in the San Joaquin River watershed and oxygen depletion in the DWSC (Lee and Jones-Lee 2003, Lehman et al. 2001). Return to Photosynthesis Water Temperature Effects on Sediment Oxygen Demand Water temperature affects the quantity and significance of SOD by affecting the rate of oxygen consumption in the sediments. The first-order reaction of SOD decay (which is similar to the BOD decay equation) is: SODt = Lo[1-e-kt]
The decay rate constant, k, is a function of temperature and would be expected to increase as temperatures increase (Chapra 1997 cited in Jassby 2005). Generally, a 10°C increase in temperature results in a doubling of the decay rate (Chapra 1997 cited in Jassby 2005). Thus, SOD would increase as water temperatures in the DWSC or San Joaquin River increased. The temperature effect on decay rates is generally assumed to be: kT = k20 • 1.05T-20
Therefore, the seasonal variation in SOD is generally strong but uncertain because there are no direct measurements. Nitrification rates are less during the winter and may cease altogether at temperatures less than 10°C (Brown 2002). Return to Sediment Oxygen Demand Flow velocity affects the following primary drivers: reaeration and residence time. Flow Velocity Effects on Reaeration and Residence Time Increased flow generally increases the flow velocity and turbulent mixing and thereby increases the reaeration rate. The increased flow velocity caused by increased flow also decreases residence time. Although reaeration and residence time are separate secondary drivers, they are closely related and therefore are discussed together in this section. Sources of flow include upstream releases from reservoirs, flow from tributaries , groundwater discharge into the San Joaquin River, agricultural and other water returns, stormwater runoff, and discharges from wastewater treatment plants and other urban and industrial sources. Diversions of water include agricultural, wetland, urban, and industrial water supply diversions; groundwater aquifer recharge; and evaporation. Diversions of water from the San Joaquin River, particularly at the head of Old River, can remove a significant amount of San Joaquin River flow from the DWSC, especially in years with high Central Valley Project and State Water Project pumping (Quinn and Tulloch 2002, Ralston and Hayes 2002). Sources of flow and flow losses are described for each river reach . Return to Reaeration Channel geometry affects the following primary drivers: reaeration and residence time. Channel Geometry Effects on Reaeration More reaeration is possible when the river channel is wide and shallow. A wider channel has more surface area for reaeration, and a shallow depth allows a larger change in DO concentration with a given rate of reaeration. Another aspect of channel geometry can effect reaeration rates: water dropping over weirs or through riffles can greatly increase the reaeration rate by increasing turbulence. This effect is more likely on the tributary streams with a substantial gradient. Channel Geometry Effects on Residence Time Channel geometry affects residence time by affecting the flow velocity. Residence time can be calculated from the flow and volume: time (days) = 0.5 • volume (af) / river flow (cfs) The channel geometry describes the biological "reactor" in which the DO dynamics of the river occur. For a given length of river, the width determines the surface area for sunlight to grow algae, macrophytes (i.e., aquatic plants), and benthic algae, and the sediment area (slightly greater than surface area). For a 1-mile segment of river, each 10 feet of width represents 1.2 acres of surface and sediment area per mile. The average depth of the river, together with the width, determines the volume of water per mile. The depth controls the effectiveness of surface reaeration in oxygenating the water column, influences the amount of photosynthesis (expressed as DO [g/m2/day]) that can take place (because of limitations on light penetration of water), and influences the extent to which SOD changes DO concentrations in the entire water column. In addition, The depth and width govern the cross-sectional area, which determines the velocity and travel time of water flowing in the reach. The channel geometry of Reaches 1 and 2 is determined primarily by natural fluvial geomorphic processes, but Reach 3 (i.e., the DWSC) was artificially created and its channel geometry is determined by channel dredging. The geometry of the DWSC controls the flow of water it and the tidal exchange and mixing in it (Jones & Stokes 2002c). The San Joaquin River channel immediately above the DWSC is 10–15 feet deep, and the channel in the DWSC is maintained to a depth of 35–40 feet (Lee and Jones-Lee 2003). As described in Residence Time [link to heading in this file], residence time and tidal mixing influence the DO decline from BOD and SOD. Therefore, its geometry is an important factor affecting the DWSC DO concentrations because it affects the residence time of oxygen-demanding materials and tidal exchange and mixing (Jones & Stokes 2002c). Return to Reaeration Algal biomass affects the following primary driver: photosynthesis. Algal Biomass Effects on Photosynthesis Algae (phytoplankton) are the primary source of photosynthetically generated DO in the study area (Lee and Jones-Lee 2003). As algal biomass increases, the amount of oxygen produced by algae through photosynthesis also increases. Studies conducted in the DWSC have shown that algal photosynthesis can add considerable amounts of oxygen to the near-surface portion of the water column. As a result, DO concentrations can be high during the day (when photosynthesis is occurring) in the near surface waters of the DWSC but low near the channel bottom where light cannot penetrate sufficiently to support it (Lehman and Ralston 2001; Jones & Stokes 2002a; Van Nieuwenhuyse 2002). Although algal biomass in the San Joaquin River can increase DO concentrations through photosynthesis, it can also decrease DO concentrations through respiration and, where slow flow rates cause algae to sink and die, through decomposition. This section discusses photosynthesis; decomposition of algae is discussed under the primary driver BOD. Algal biomass in the study area is derived from algae that enter the study area from upstream sources (imported BOD) and algae that are produced in each reach. In the study area, algal growth is primarily a function of the temperature of water, the amount of available light, and the amount of available algal nutrients (Thuman et al. 2004). In addition, algal biomass is also affected by the time available for algae to grow in the study area. For example, increased algal biomass in the DWSC will generally occur during periods of increased residence times (Quinn and Tulloch 2002). Imported algal biomass is the algal biomass that is contributed to the San Joaquin River in water from tributaries, return flows, and other flow sources. The imported algae provide the seed for algal growth and generally increasing concentrations in the river. Turbidity affects the following primary driver: photosynthesis. Turbidity Effects on Photosynthesis Photosynthesis and the subsequent production of oxygen require the presence of sunlight (Tchobanoglous and Schroeder 1985). Once sunlight reaches the water surface, the clarity of the water determines the depth to which light travels into the water column. The euphotic zone is the portion of the water column receiving sufficient light for photosynthesis. Suspended particles in the water column can reduce the clarity of the water, decreasing the depth of the euphotic zone and decreasing the amount of sunlight reaching that zone. The sources of the suspended sediment in the San Joaquin River are not well documented. However, potential sources include scour of mud from the channel bottom, erosion of soil, and input of other particulate matter from stormwater runoff, tributaries, and other flow sources. In addition to turbidity, colored water released from managed wetlands in the Mud and Salt Slough watershed has been shown, at times, to contribute sufficient color to the San Joaquin River and DWSC to reduce light penetration sufficiently to affect the rate of algal photosynthesis (Lee and Jones-Lee 2003). Although algae in the San Joaquin River are mixed and grow at the average light level in the river, in the DWSC, the algae are not in the surface lighted layer (euphotic zone) for enough of the day to increase in the net biomass or DO production. Reductions in the availability of light associated with increasing turbidity can also reduce the lifespan of algae established in the affected water column (Lee and Jones-Lee 2000). Periods of the highest turbidity in the study area are likely experienced during February through May, when flows are highest and particulate materials are eroded and flushed downstream. One study found that in 2001, turbidity values in the DWSC declined from June to October (Jones & Stokes 2002a). Turbidity values in the San Joaquin River from Vernalis to Channel Point were similar to those measured in the DWSC and experienced a similar decline during the summer (Jones & Stokes 2002a). Nutrients affect the following primary driver: photosynthesis. Nutrient Effects on Photosynthesis Algae use nutrients during photosynthesis. Nitrogen and phosphorus are the primary nutrients required for algal growth, as well as carbon obtained from dissolved CO2 in the water column. Conversely, a surplus of nutrients can lead to eutrophication and excessive algae growth, which can reduce DO concentrations. Nutrients are contributed to the San Joaquin River by upstream sources, as well as local stormwater runoff and wastewater discharges (Lee and Jones-Lee 2003). Nutrients in the study area are derived from soil, fertilizers, animal waste from dairies and other livestock-related operations, and discharges to the river from wastewater treatment facilities (Leland et al. 2001). Ammonia inputs to the San Joaquin River from the Stockton RWCF have generally been lowest during the summer (i.e., less than 10 mg/L) and highest (i.e., 25 mg/L ammonia-N) during the winter (Jones & Stokes 2002a). One indication that human activities may substantially contribute nutrients to the San Joaquin River is that nitrogen isotopes in the river (upstream of Vernalis) appear to be derived from animal waste and sewage discharges (Kratzer et al. 2004). This nitrogen isotope analysis also indicated that soil nitrogen and/or fertilizer are the most important nitrogen sources in tributaries to the San Joaquin River (Kratzer et al. 2004). Imported Biochemical Oxygen Demand Imported BOD affects the following primary driver: BOD. Imported Biochemical Oxygen Demand Effects on Biochemical Oxygen Demand Imported BOD concentration is the concentration of BOD-generating substances (e.g., algal biomass) that are contributed to the San Joaquin River in water from upstream reaches, tributaries, agricultural return flows, and other flow sources. Sources of imported BOD are described in the reach-specific discussions. Imported BOD concentrations affect the BOD concentrations in the San Joaquin River and the DWSC. When the imported BOD concentrations are greater than the existing BOD concentrations (i.e., in the San Joaquin River or DWSC), the BOD concentrations in the San Joaquin River or DWSC increase. Conversely, when the imported BOD concentrations are lower than the existing BOD concentrations, the BOD concentrations in the San Joaquin River or DWSC decrease. The effects of imported BOD on BOD concentrations downstream are influenced by:
Carbonaceous Biochemical Oxygen Demand CBOD affects the following primary driver: BOD. Definition CBOD results when oxygen is consumed by microorganisms in converting organic material into CO2, H20, nutrients, energy and new cells (Lee and Jones-Lee 2003). Algal cells contain organic chemicals that will consume oxygen during decomposition. The following chemical equation provides estimates of the quantity of CBOD that will be produced during the decomposition of an algal cell (Litton 2003). CO2 + H2O + Nutrients (in the presence of light) —> CH2O [new algal cells] + O2 CBOD is made up of both algal and nonalgal biomass. Carbonaceous Biochemical Oxygen Demand Effects on Biochemical Oxygen Demand Algal Biomass The current body of information indicates that algae and algal detritus are the primary sources of BOD in the DWSC (Lee and Jones-Lee 2003). Each 1 mg/L of algae will yield a theoretical total oxygen demand of 1.2 mg/L, which represents the sum of CBOD and NBOD. The CBOD from the decomposition of 1 mg/L of algal biomass is 0.9 mg/L, or 75% of the total oxygen demand, while the NBOD is 0.3 mg/L or 25% of the total oxygen demand (Litton 2003). The algae pigments chlorophyll a and pheophytin are used to represent the concentrations of living and dead algae, respectively, in a water column. Therefore, the pheophytin concentrations indicate the quantity of dead algae that might be a source of CBOD. However, the chlorophyll a and pheophytin content of algal biomass is somewhat variable. For more information about algal biomass, see Algal Biomass. Nonalgal Biomass Although algae are believed to be the principle source of organic matter in the San Joaquin River, there are other sources as well. Limited information is available on these nonalgal biomass sources. However, potential sources include:
Total organic carbon is a chemical measurement technique that indicates all oxidizable carbon in a sample. Some of this material is only partially decayed by microorganisms and does not contribute to BOD. Salt Slough, Mud Slough, and the Tuolumne and Stanislaus Rivers are the most significant tributary sources of dissolved organic carbon to the San Joaquin River and accounted for as much as 2445% of the 2000 and 2001 dissolved organic carbon loads in the San Joaquin River at Vernalis (Kratzer et al. 2004). Dissolved organic carbon concentrations, measured at several locations in the San Joaquin River upstream of Vernalis, were relatively constant from June through September in 2000 and 2001 (Kratzer et al. 2004). Organic carbon loads in the San Joaquin River increased in October and November because of wetland drainage into Mud Slough and reservoir releases on the Merced River (Kratzer et al. 2004). One study suggests that, in the San Joaquin River and in the DWSC, live and detrital algae comprised, on average, only 2040% of the total organic carbon (). Therefore, nonalgal organic matter loads may be an important source of organic carbon and BOD. Nitrogenous Biochemical Oxygen Demand NBOD affects the following primary driver: BOD. Direct inputs of ammonia to the San Joaquin River include discharges from wastewater treatment plants and wastewater from dairies and other animal husbandryrelated operations (Lee and Jones-Lee 2000, Litton 2003). WWWTP dischargers to tributaries of the San Joaquin River include Newman WWTP, Turlock WWTP, and Los Banos WWTP. The City of Modesto WWTP (during the winter) and the City of Manteca WWTP discharge directly to the San Joaquin River (Quinn and Tulloch 2002). The City of Stockton discharge is the largest source of wastewater BOD and ammonia to the San Joaquin River upstream of the DWSC. The contribution of ammonia concentrations to oxygen demand in the DWSC was identified as early as the late 1960s (McCarty 1969 in Foe et al. 2002, Lehman 2003). Particulate Biochemical Oxygen Demand Concentration Particulate BOD concentration affects the following primary driver: SOD. Particulate BOD Effects on SOD Algae biomass represents a significant source of BOD when it is suspended in the water column. However, when the algae biomass settles out of the water column, this biomass represents a source of SOD. Particulate BOD in the water column that may exert BOD also represents a source of SOD if it settles out of the water column. Residence time affects SOD concentrations because particulate BOD, such as dead algae, settles from the water column and contributes to SOD. As residence time increases, the amount of particulate BOD that settles out also increases, which in turn contributes to an increase in SOD. Iron and Sulfate Compound Concentration The concentration of iron and sulfate compounds affects the following primary driver: SOD. Iron and sulfate compound concentrations contribute to SOD through a series of chemical processes in sediments under anaerobic conditions (i.e., absence of oxygen). Organics material in the water column settle out and contribute to the biotic SOD. As this process consumes DO available in the sediment, it allows other reduction reactions among iron and sulfate compounds. The resultant ferrous iron and sulfide constituents in the sediments are then available to dissolve or mix in the water column and take up DO in abiotic reactions (Lee and Jones-Lee 2000). |